{kind=link}

Thursday, December 29, 2016
Early Indo-European migrations map
Wikipedia has a new animated gif of early Indo-European migrations (available at various resolutions here). It's pretty good overall, but very speculative and potentially erroneous in parts. For instance, my understanding is that the Vedic Aryans did not emerge from BMAC per se, as the map suggests, but rather from a post-BMAC phenomenon heavily influenced by steppe pastoralists. Hi-res ancient DNA from BMAC and post-BMAC sites should be able to resolve this issue.
 As far as I know, BMAC remains were being tested at Harvard earlier this year, but the year is almost out, and nothing has been published. So either David Reich and co. are keeping the results for a new paper on the Indo-European homeland question, or they couldn't get any usable data from the samples. Keep in mind that only 30-40% of the ancient remains that are tested at Harvard are successfully genotyped. I can imagine that the success rate for samples from arid locations, like former BMAC sites in Turkmenistan, is even lower.
Update 31/12/2016: Commentator Tapatuevik Kaarmkyno points us to an article from earlier this year at NIH Record featuring this quote from David Reich:"We’ve sequenced more than 1,000 samples in our own lab — there’s not enough time to publish". That's probably why the second half of 2016 was so agonizingly slow. Next year should be awesome.
See also...
Maybe first direct hints of Yamnaya-related gene flow into South Central Asia
As far as I know, BMAC remains were being tested at Harvard earlier this year, but the year is almost out, and nothing has been published. So either David Reich and co. are keeping the results for a new paper on the Indo-European homeland question, or they couldn't get any usable data from the samples. Keep in mind that only 30-40% of the ancient remains that are tested at Harvard are successfully genotyped. I can imagine that the success rate for samples from arid locations, like former BMAC sites in Turkmenistan, is even lower.
Update 31/12/2016: Commentator Tapatuevik Kaarmkyno points us to an article from earlier this year at NIH Record featuring this quote from David Reich:"We’ve sequenced more than 1,000 samples in our own lab — there’s not enough time to publish". That's probably why the second half of 2016 was so agonizingly slow. Next year should be awesome.
See also...
Maybe first direct hints of Yamnaya-related gene flow into South Central Asia
Saturday, December 24, 2016
Do it right or don't do it at all
At least a couple of academic teams working on the population history of Europe need to read this PDF and take it to heart.
This article reviews scientific publications that have attempted to use genetic and genomic data in order to investigate European migrations between the fourth and ninth centuries. It considers early single-locus studies that used mtDNA and y-chromosome data. These studies were successful in formulating hypotheses concerning migration and heterogeneity, primarily between the Continent and the British Isles and Iceland, but could only examine a small portion of the entire genetic inheritance. The article continues with a presentation of more recent genome-wide studies. In particular, it evaluates the problems of using modern genomic data to understand past migratory processes, arguing that modern DNA is a problematic source for understanding population histories of the past fifteen hundred years and urges the sequencing and analysis of ancient DNA. It also presents some of the problems of research teams that did not include archaeologists and historians as integral participants in the planning, collection, and evaluation of data. It concludes with a brief outline of the authors’ current project that examines migration between Pannonia and Italy in the sixth century. ... This work is necessarily interdisciplinary, something notably lacking in the majority of genetic studies. The Ralph and Coop study, while highly rigorous at the level of the population genetic analysis, included no historians or archaeologists, and the only historical literature cited, presumably to »identify« the Hunnic contribution to European population, was a general history of Europe, [25] a survey of Slavic history, [26] and two articles in the New Cambridge Medieval History. [27] The Busby et al. study also included no historians or archaeologists on its team, and the only historical literature cited was a Penguin History of the World, Peter Heather’s survey of the Early Middle Ages, and a survey of Muslims in Italy. [28] Unlike these studies, designed and executed exclusively by geneticists who then look through a few general historical handbooks to try to find stories that might explain their data, historians and archaeologists are integrated from the start in our project.Patrick J. Geary and Krishna Veeramah, Mapping European Population Movement through Genomic Research [PDF download], Medieval Worlds, No. 4, 201, 65-78
Sunday, December 18, 2016
So how many of you really read the preprints at bioRxiv?
There's a new preprint at bioRxiv about the spread of the plague across much of Eurasia via the expansions of Early Bronze Age steppe pastoralists. It's been up for a few days now. Judging by the number of tweets on the preprint, a lot of people have read it. Or have they?
The manuscript contains a glaring error, a couple times over, in which the authors mix up the Afanasievo Culture with the Andronovo Culture. That's OK, this is why we have preprints and bioRxiv. I just found it somewhat amusing to see one lonely voice pointing this out under the rash of tweets recommending the paper.
 Update 20/12/2016: A corrected version of the preprint has just been posted at bioRxiv. See here.
See also...
Mobile and then some
Update 20/12/2016: A corrected version of the preprint has just been posted at bioRxiv. See here.
See also...
Mobile and then some

Saturday, December 17, 2016
The Metal Age invader that never was
FTDNA has launched a new "Ancient Origins" analysis, and I've been asked a few times now what I think of the "Metal Age invader" component in this test. Essentially, I share the view of one of the commentators in the comments here.

See also... Massive migration from the steppe is a source for Indo-European languages in Europe (Haak et al. 2015 preprint).

Wolfgang Haak et al., Massive migration from the steppe is a source for Indo-European languages in Europe, bioRxiv, Posted February 10, 2015, doi: http://dx.doi.org/10.1101/013433
See also... The Metal Age invader that never was #2 Who's your (proto) daddy Western Europeans?
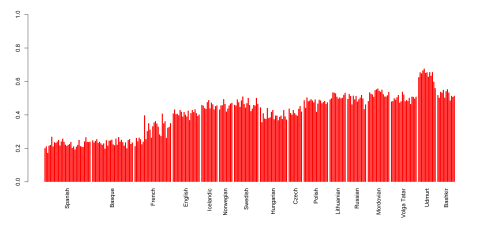
FTDNA now models Europeans as a mix of hunter-gatherers (Loschbour, La Brana 1 and Motala), early European farmers (Stuttgart, Iceman, LBK) and so called "Metal Age invaders" which are based on Corded Ware and Yamnaya samples - so the latter is what we more typically call "steppe admixture". (Plus they add non-European admixture in case someone has it very evidently.) This model looks crude for our present-day standards, and their results look very different from what I've seen here in Davidski's analyses or in other ancient-DNA calculators on Gedmatch, as their inferred proportions of steppe admixture are much lower, about half of that inferred by others. And moreover to me it looks completely wrong that FTDNA suggests that I (1/4 Italian, strong southwest European component) have more Metal Age invader admixture than my fully East Prussian grandmother.Indeed, any Early Bronze Age Steppe-inspired component should peak at 40-50% amongst Northern and Eastern Euros. This apparently doesn't. Why? No idea. I know better than almost anyone else that there's no such thing as a perfect ancestry test, and FTDNA has every right to offer an experimental analysis to its clients, so let's leave it at that. But if you want to see what an ADMIXTURE test with a half-decent Yamnaya component looks like, then check out my attempt HERE. These are graphs from the linked blog entry showing inferred levels of Yamnaya-related ancestry proportions for Europeans and West Asians.
See also... Massive migration from the steppe is a source for Indo-European languages in Europe (Haak et al. 2015 preprint).
Wolfgang Haak et al., Massive migration from the steppe is a source for Indo-European languages in Europe, bioRxiv, Posted February 10, 2015, doi: http://dx.doi.org/10.1101/013433
See also... The Metal Age invader that never was #2 Who's your (proto) daddy Western Europeans?
Mobile and then some
Update 07/12/2018: Europe's ancient proto-cities may have been ravaged by the plague
...
I've now read the new Valtuena et al. prerprint a couple of times, and even though the plague angle is interesting, what really sticks out for me is how incredibly mobile our steppe ancestors were.
It's been obvious for a while now that during the Early Bronze Age there was a massive expansion from the Pontic-Caspian Steppe in all directions, as far as the Atlantic shores in the west and the Altai region in the east. However, considering the phylogeny of Yersinia pestis genomes in Valtuena et al., it looks like the Yamnaya and/or closely related peoples also regularly roved across the entire length of the Eurasian Steppe.
The oldest and phylogenetically most basal sample of Yersinia pestis to date comes from an Afanasievo skeleton from the Altai region dated to 4836-4625 YBP. Hot on its heels, the next oldest is a sample derived from the Afanasievo strain, but from a Corded Ware skeleton from the East Baltic dated to 4571-4422 BP. The two burial sites are separated by something like 3,000 kilometers.
It's unlikely that Yersinia pestis spread from the Pontic-Caspian Steppe, say, with the Yamnaya, both to the Altai region and the East Baltic, because it hasn't yet been found in any of the Yamnaya remains. It's also generally accepted that Yersinia pestis is indeed from Asia. So it must have been picked up by migrants from the Pontic-Caspian Steppe somewhere in Asia, probably the Altai region, whose descendants then returned to Europe with the bug, and may have taken part in the Corded Ware expansion.
But of course it doesn't end there, because it appears that the descendants or close relatives of the Baltic Corded Ware people then moved back to Asia, because a sample of Yersinia pestis derived from the Corded Ware strain is found in an Andronovo skeleton from the Altai region dated to 3694-3575 BP. Note also that the genome-wide genetic structure of most of the Andronovo individuals sampled to date is Corded Ware-like, in fact more so than Yamnaya-like. In other words, ancient human DNA also shows a very close relationship between Andronovo and the older Corded Ware.
Why were these people roving around so much? I know they were pastoralists and that the Eurasian Steppe became progressively drier during the Bronze Age, so it may have been necessary to move wherever suitable pastures were available. But still, it seems rather ridiculous that they spent so much energy on long range intercontinental travel when, by all accounts, all they had were wagons pulled by oxen.
Citation...
Valtuena et al., The Stone Age Plague: 1000 years of Persistence in Eurasia, bioRxiv, Posted December 15, 2016. doi: https://doi.org/10.1101/094243
See also...
So how many of you really read the preprints at bioRxiv?
Bronze Age dope dealers
Thursday, December 15, 2016
More on the stone age plague
Update 07/12/2018: Europe's ancient proto-cities may have been ravaged by the plague
...
Good stuff at bioRxiv:
 Valtuena et al., The Stone Age Plague: 1000 years of Persistence in Eurasia, bioRxiv, Posted December 15, 2016. doi: https://doi.org/10.1101/094243
See also...
Plague germs may have facilitated Bronze Age expansions from the steppe
Mobile and then some
So how many of you really read the preprints at bioRxiv?
Valtuena et al., The Stone Age Plague: 1000 years of Persistence in Eurasia, bioRxiv, Posted December 15, 2016. doi: https://doi.org/10.1101/094243
See also...
Plague germs may have facilitated Bronze Age expansions from the steppe
Mobile and then some
So how many of you really read the preprints at bioRxiv?
Abstract: Molecular signatures of Yersinia pestis were recently identified in prehistoric Eurasian individuals, thus suggesting Y. pestis might have caused some form of plague in humans prior to the first historically documented pandemic. Here, we present four new Y. pestis genomes from the European Late Neolithic and Bronze Age (LNBA) dating from 4,500 to 3,700 BP. We show that all currently investigated LNBA strains form a single genetic clade in the Y. pestis phylogeny that appears to be extinct today. Interpreting our data within the context of recent ancient human genomic evidence, which suggests an increase in human mobility during the LNBA, we propose a possible scenario for the spread of Y. pestis during the LNBA: Y. pestis may have entered Europe from Central Eurasia during an expansion of steppe pastoralists, possibly persisted within Europe until the mid Bronze Age, and moved back towards Central Eurasia in subsequent human population movements. ... The first indication of plague in Europe is found in the Baltic region and coincides with the time of the arrival of the steppe component (Allentoft et al., 2015). The two Late Neolithic Y. pestis genomes from the Baltic in this study were reconstructed from individuals associated with the Corded Ware Complex (Gyvakarai1 and KunilaII). The Baltic Y. pestis genomes are genetically derived from the strain that was found in the ‘Andronovo Complex’ from the Altai region [my note: I think they mean Afanasievo], suggesting that the disease might have spread with steppe pastoralists from Central Eurasia to Eastern and Central Europe during their massive range expansion. The younger Late Neolithic Y. pestis genomes from Southern Germany are genetically derived from the Baltic strains and are found in individuals associated with the Bell Beaker Complex. Previous analysis have shown that Bell Beaker individuals from Germany also carry ‘steppe ancestry (Allentoft et al., 2015; Haak et al., 2015). This suggests that Y. pestis may have been spread further southwestwards analogous to the human steppe component. The youngest of the LNBA Y. pestis genomes (RISE505), found also in the Altai region, descends from the Central European strains, and thus suggest a spread back into the eastern steppes.

Thursday, December 8, 2016
Middle Ages: rotten time to be a spotted horse
Scientific Reports has a new paper on the history of coat patterns in domesticated horses and the domestication process itself. Emphasis is mine:
Horses have been valued for their diversity of coat colour since prehistoric times; this is especially the case since their domestication in the Caspian steppe in ~3,500 BC. Although we can assume that human preferences were not constant, we have only anecdotal information about how domestic horses were influenced by humans. Our results from genotype analyses show a significant increase in spotted coats in early domestic horses (Copper Age to Iron Age). In contrast, medieval horses carried significantly fewer alleles for these phenotypes, whereas solid phenotypes (i.e., chestnut) became dominant. This shift may have been supported because of (i) pleiotropic disadvantages, (ii) a reduced need to separate domestic horses from their wild counterparts, (iii) a lower religious prestige, or (iv) novel developments in weaponry. These scenarios may have acted alone or in combination. However, the dominance of chestnut is a remarkable feature of the medieval horse population. ... In addition, we discovered that tobiano spotting, which only occurs in domestic horses and had thus far only been detected after 1500 BC, was present in the Eneolithic/Copper Age (Kazakhstan, cal. 3654–3630 BC, and Germany, cal. 3368–3101 BC). Similar to chestnut and sabino spotting, the tobiano phenotype appears to arise shortly after domestication, which is assumed to have started approximately 4000–3500 BC in the Ponto-Caspian steppe region (modern day Kazakhstan and Ukraine) [17]. ... Moreover, the detection of the tobiano phenotype in two Eneolithic domestic horses from distant regions has important implications regarding the origins of horse domestication [17] and their subsequent distribution. It also supports previous claims that the emergence of domestic horses in Central Europe at the end of the fourth millennium BC was facilitated by introduced horses from the Ponto-Caspian steppe [19].Wutke, S. et al. Spotted phenotypes in horses lost attractiveness in the Middle Ages. Sci. Rep. 6, 38548; doi: 10.1038/srep38548 (2016). See also... Sintashta and Scythian horses came from Hyperborea
Wednesday, November 30, 2016
Five foot Philistines
Vilified in biblical texts and even ridiculed by Iggy Pop, the Iron Age pagan seafarers known as the Philistines were basically the East Mediterranean version of the Vikings, except apparently much smaller:
Relatively short average heights for people buried at Ashkelon — about 5 feet, 1 inch for men and 4 feet, 10 inches for women — also fit a scenario of biological stress, Fox said. Short stature and minimal height differences between men and women occur with population-wide stresses such as malnutrition, she said. ... The Philistines were a famously combative crowd. Archaeologist Eric Meyers of Duke University, who was not a member of the Ashkelon team, wondered if at least some of those buried at Ashkelon had been killed in battles or fights. But no head injuries or other skeletal signs of violent encounters appeared among the dead at Ashkelon, Fox said. Neither did any skeletons contain evidence of tumors or cancers. If DNA can be extracted from the Ashkelon skeletons, scientists may get a glimpse of where the Philistines originally came from. Evolutionary geneticist Johannes Krause of the Max Planck Institute for the Science of Human History in Jena, Germany, is currently directing efforts to retrieve genetic sequences from the Ashkelon bones.Source: Ancient cemetery provides peek into Philistines’ lives, health, ScienceNews, November 22, 2016 See also... Evidence of European ancestry in the Philistines
Saturday, November 26, 2016
Big deal of 2016: the territory of present-day Iran cannot be the Indo-European homeland
It's been a pretty big year. Not as big as many of us had hoped for, but there's still a few more weeks till 2017, so who knows what will happen?
In any case, for me 2016 will be the year in which we finally saw hard data - courtesy of Lazaridis et al. and Broushaki at al. - that crossed off the territory of present-day Iran from the list of potential Proto-Indo-European (PIE) homelands.
Now, it's true, Lazaridis et al. and Broushaki et al. were somewhat vague in what their data meant in this context, and indeed, both sets of authors left open the possibility that what is now Iran might prove to be the PIE homeland. However, their data left no doubt: look elsewhere for the PIE Urheimat.
Why? Here it is, in point form, as simply as I can put it:
- the Mesolithic and Neolithic peoples of the Zagros range and South Caspian region were highly distinct both in terms of genome-wide genetic structure and uniparental markers, and clearly only contributed meaningful gene flow to South Asia, not to Europe or Anatolia - after the descendants of Neolithic farmers from the Zagros range, or at least their very close relatives, migrated to South Asia, the territory of present-day Iran, as per the data in Lazaridis et al. and Broushaki et al., saw waves of migrations from the west and north that dramatically shifted the population structure of the region, bringing it closer in this respect to the Levant and Europe - most of the Neolithic samples from Iran in Lazaridis et al. and Broushaki et al. came from near the proto-Elamite homeland in the Kor River basin in central Fars, which strongly suggests that their close relatives who streamed into South Asia were not Indo-Europeans, but rather speakers of languages closely related to Elamite - even though Lazaridis et al. successfully modeled Early/Middle Bronze Age steppe populations, including Yamnaya, as largely of Iran Chalcolithic origin, this appeared to be a coincidence, because the Chalcolithic samples from Iran showed fairly typical South Caspian uniparental markers, such as Y-chromosome haplogroups J and G1 and mitochondrial (mtDNA) haplogroup U7, which are conspicuous by their absence from an exceptionally wide range of Bronze Age steppe samples.I do realize that many readers won't accept these arguments for emotional reasons, because the PIE homeland debate is an emotional one for a lot of people. Nevertheless, if you decide to argue with me in the comments, make sure you have a coherent argument. Actually, the territory of present-day Iran has never been a serious contender in the PIE homeland debate. This is something that many Iranian scholars will freely admit. Not only was central Fars the proto-Elamite homeland, but much of what is now western Iran was the stomping ground of the Hurrians. The Indo-Europeans, in the form of Indo-Iranians such as the Medes, only got there late in the game, in all likelihood from the Eurasian steppes. But a lot of newbies to the PIE debate don't know this, or they don't want to know it. I guess to them Iran makes sense because it's in between Europe and South Asia? Or maybe it's the fact that the word Iran is kind of similar to the word Aryan? No idea, really, but like I say, it's now all over. See also... A homeland, but not the homeland Zarathushtra and his steppe posse The story of mtDNA haplogroup U7
Thursday, November 24, 2016
Multiple Holocene Eurasian migrations into north central Africa
The AJHG has a new open access paper on the population history of Chad. It lays out a reasonable hypothesis for the sources and timing of Eurasian gene flow into north central Africa. Just wondering, though, if the authors considered the possibility that R1b-V88 may have expanded into Africa from Iberia? After all, the oldest instance of a probable R1b-V88 to date is from an Iberian Early Neolithic sample (ID I0410, Els Trocs, Spain, Haak et al. 2015). But I don't have a strong opinion on the issue at the moment. From the paper (emphasis is mine):
Understanding human genetic diversity in Africa is important for interpreting the evolution of all humans, yet vast regions in Africa, such as Chad, remain genetically poorly investigated. Here, we use genotype data from 480 samples from Chad, the Near East, and southern Europe, as well as whole-genome sequencing from 19 of them, to show that many populations today derive their genomes from ancient African-Eurasian admixtures. We found evidence of early Eurasian backflow to Africa in people speaking the unclassified isolate Laal language in southern Chad and estimate from linkage-disequilibrium decay that this occurred 4,750–7,200 years ago. It brought to Africa a Y chromosome lineage (R1b-V88) whose closest relatives are widespread in present-day Eurasia; we estimate from sequence data that the Chad R1b-V88 Y chromosomes coalesced 5,700–7,300 years ago. This migration could thus have originated among Near Eastern farmers during the African Humid Period. We also found that the previously documented Eurasian backflow into Africa, which occurred ∼3,000 years ago and was thought to be mostly limited to East Africa, had a more westward impact affecting populations in northern Chad, such as the Toubou, who have 20%–30% Eurasian ancestry today. We observed a decline in heterozygosity in admixed Africans and found that the Eurasian admixture can bias inferences on their coalescent history and confound genetic signals from adaptation and archaic introgression. ... It is important to note that in this work we inevitably invoke Occam’s razor to support the simplest model consistent with our data; the history of the populations studied here, including the time and sources of the Eurasian admixture in Africa, could be more complex. aDNA from Chad and neighboring regions remains a challenge given the poor DNA preservation in hot climates, but future successful efforts in aDNA research could provide additional insights and reveal additional complexities not considered by the modern-DNA-based models favored here.Haber at al., Chad Genetic Diversity Reveals an African History Marked by Multiple Holocene Eurasian Migrations, AJHG, Published Online: November 23, 2016, DOI: http://dx.doi.org/10.1016/j.ajhg.2016.10.012 show See also... R1b-V88: out of the Balkans and into Africa?
Saturday, November 12, 2016
Days of high adventure
I've redesigned and streamlined my Principal Component Analysis (PCA) plot of West Eurasia in anticipation of the arrival of many more ancient samples. Rumor has it we'll not only get stuff from the Balkans, but also finally from the steppes north of the Black Sea and South Asia.
I'd say my new plot does a better job of highlighting relationships between the different prehistoric groups and population shifts across space and time. The datasheet is available here.
 It should be pretty clear from this plot how the modern-day European gene pool came about. So I don't expect any major surprises when the new samples come in. Nevertheless, the wait is killing me, and many others I'm sure.
Update 13/11/2017: A year on, I've acquired many new samples and, as a result, streamlined the PCA some more, see...
Who's your (proto) daddy Western Europeans?
It should be pretty clear from this plot how the modern-day European gene pool came about. So I don't expect any major surprises when the new samples come in. Nevertheless, the wait is killing me, and many others I'm sure.
Update 13/11/2017: A year on, I've acquired many new samples and, as a result, streamlined the PCA some more, see...
Who's your (proto) daddy Western Europeans?

Monday, November 7, 2016
Cryptic post-OOA African ancestry in Eurasians (?)
The Max Planck Institute for the Science of Human History is holding the Human Dispersals in the Late Pleistocene - Interdisciplinary Approaches Towards Understanding the Worldwide Expansion of Homo sapiens conference this week. The conference web page and abstract book are here.
The abstract below from Stephan Schiffels is very interesting. It reminds me of some of the discussions that we've had in the comments about potential African ancestry in some Eurasians that might be hiding due to a lack of relevant ancient African samples (for instance, see here).
When and how modern humans left the African continent is still a debated question. Recently, three projects have analysed new genetic data from modern populations in Papua New Guinea and Australia, which has provided new insights on this topic. I will present analyses from one of these publications (Malaspinas et al. 2016), and compare results with findings from the two other projects (Mallick et al. 2016, Pagani et al. 2016). Here, we used MSMC2, a novel computational framework to analyse the distribution of times to the most recent common ancestor along multiple sequences. We find that all non-African populations that we analysed, including Australians, experienced a very similar population bottleneck in the past, consistent with only one out-of-Africa migration for all extant non-African populations. At the same time, we find evidence that some African populations are more distantly related to Australians than to Eurasian populations, and we show that this result is robust to haplotype phasing errors and archaic introgression. We interpret our result as evidence for gene flow between some Africans and Eurasians after the initial split, which is also consistent with results from other population genetic methods. Our analysis suggests that in order to understand human dispersal out of Africa, we need to better understand ancient population substructure within Africa, which is an important direction for future research.
Stephan Schiffels, Analysing Australian genomes to learn about early modern human dispersal out of Africa, Human Dispersals in the Late Pleistocene - Interdisciplinary Approaches Towards Understanding the Worldwide Expansion of Homo sapiens 2016 conference abstract
See also...
Two pronged AMH colonization of Eurasia. Or not
Friday, November 4, 2016
Caste is in the genes
I've built up a pretty good record over the past couple of years of pre-empting results that eventually show up in high end scientific journals.
Here's another major finding that you'll see at some point in Nature or Science: in present-day India upper caste membership is strongly associated with Eneolithic/Bronze Age steppe-related ancestry, while low caste membership and Dravidian languages are strongly associated with indigenous South Asian and Zagros Neolithic-related ancestry.
As far as I can see, with my somewhat limited dataset, the best way to show this is by plotting D-stats of the form D(Outgroup;Samara_Eneolithic)(Onge;X) versus D(Outgroup;Iran_Neolithic)(Onge;X). The relevant datasheet is available here.
 Like I say, my dataset is still not great for this type of analysis, but the results are very striking and I have no doubt that they will be confirmed with more comprehensive sampling of Indian populations.
If they are confirmed, this will probably mean three things: 1) the caste system was introduced into the Indian subcontinent from the Eurasian steppe after the demise of the Chalcolithic Harappan civilization, 2) Zagros Neolithic farmers and closely related Neolithic/Chalcolithic populations of South Asia were not Indo-European-speaking, 3) like the caste system, Indo-Aryan (and thus Indo-European) languages were introduced into the Indian subcontinent from the Eurasian steppe after the Chalcolithic.
See also...
On the doorstep of India
Indian confirmation bias
The Out-of-India Theory (OIT) challenge: can we hear a viable argument for once?
Like I say, my dataset is still not great for this type of analysis, but the results are very striking and I have no doubt that they will be confirmed with more comprehensive sampling of Indian populations.
If they are confirmed, this will probably mean three things: 1) the caste system was introduced into the Indian subcontinent from the Eurasian steppe after the demise of the Chalcolithic Harappan civilization, 2) Zagros Neolithic farmers and closely related Neolithic/Chalcolithic populations of South Asia were not Indo-European-speaking, 3) like the caste system, Indo-Aryan (and thus Indo-European) languages were introduced into the Indian subcontinent from the Eurasian steppe after the Chalcolithic.
See also...
On the doorstep of India
Indian confirmation bias
The Out-of-India Theory (OIT) challenge: can we hear a viable argument for once?

Friday, October 28, 2016
Steppe boys, farmer girls
I updated my table of Steppe_EMBA mitochondrial (mtDNA) haplogroups with data from Wilde 2014, Broushaki et al. 2016, Sokolov et al. 2016, Jones et al. 2015, and extra data from Lazaridis et al. 2016. The accompanying map is from Wilde 2014 and shows sampling locations of many of the individuals in the spreadsheet.

 Last time I looked at this stuff when attempting to find a link between Steppe_EMBA and prehistoric Iran, I couldn't spot anything meaningful (see here). At least now I can see a few shared haplogroups. Some thoughts:
Last time I looked at this stuff when attempting to find a link between Steppe_EMBA and prehistoric Iran, I couldn't spot anything meaningful (see here). At least now I can see a few shared haplogroups. Some thoughts:

- Despite the small sample set from Neolithic Bulgaria and western Ukraine, two out of the four haplogroups are shared with Steppe_EMBA, including the fairly specific H2a2 - Steppe_EMBA potentially shares very deep ancestry with Iran_Neolithic, via basal clades such as T2c and possibly X2; these markers could have arrived on the steppe in any number of ways, and indeed may have been sitting there since the Mesolithic - But Iran_Chalcolithic is still a horrible match with Steppe_EMBA, and Maikop no betterNeedless to say, I'm looking forward to those new Neolithic Balkan samples from the upcoming Mathieson et al. paper (see here). I'm pretty sure now that at least some Steppe_EMBA groups will show admixture from Neolithic or Chalcolithic populations from the eastern Balkans. See also... Mixed marriages on the early Eneolithic steppe Modeling Steppe_EMBA Male-dominated conquest of Europe by Bronze Age steppe pastoralists The story of mtDNA haplogroup U7
Thursday, October 27, 2016
Strong mitogenomic continuity on the Armenian Plateau since the early Neolithic
Below is another abstract from the upcoming 6th DNA Polymorphisms in Human Populations conference in Paris. It was added to the abstract book after I first blogged about the conference here.
The authors are probably first testing mtDNA to check which of the samples have enough DNA for full genome sequencing or enrichment capture of genome-wide SNPs. So the fact that they managed to sequence so many mitogenomes means that we might soon see genome-wide data for most of the same samples.
The results make sense considering the genetic structure of present-day Armenians (see here). Indeed, I suspect that early Neolithic farmers from the southern Caucasus will come out looking very similar overall to present-day Armenians.
If so, this will probably be hailed by many as evidence supporting the Armenian Plateau Indo-European homeland theory. However, as I've already shown, it's very likely that the Armenian Plateau was affected by population movements from the Eastern European steppe during the Bronze Age which may have introduced Indo-European languages to the region (see here).
The origin of the Armenian people is heavily debated among historians and archaeologists. Despite a long history and vast archaeological records in Armenia, it has proven very challenging to infer the demographic events that led to the formation of Armenians as a distinct ethno-cultural group. To obtain a detailed understanding of the demographic events in Armenia across millennia, we study complete mitochondrial genomes from 49 ancient individuals covering 7800 years and compare them with that of modern Armenians (n=206) and seven neighboring populations (n=482). In this context, the lowest genetic distance was observed between the modern and ancient Armenians and this was also reflected in network analyses and discriminant analysis of principal components (dapc) showing genetic proximity between the ancient individuals and modern Armenians. We used Approximate Bayesian Computation (ABC) to test five different demographic scenarios of the Armenian population, and the simulations favored a model where both ancient and modern Armenians derive from the same source population. We conclude that there is a strong signal of continuity in the maternal Armenian gene pool during the last 7800 years.Margaryan et al., 7800 years of Mitochondrial genetic continuity in Armenia, 6th DNA Polymorphisms in Human Populations, Talk Workhsop Genomic Demography, Musee de l’Homme, Paris, 7-10 December, 2016 Update 29/06/2017: The paper has just been published. It only has a few samples from before the Middle Bronze Age (MBA) and these don't match closely the mtDNA lineages from the Eneolithic/Bronze Age steppe, which is interesting and potentially important for the Indo-European homeland debate. See here.
Thursday, October 20, 2016
Sibero-Scythian idol's "racial realignment"
From The Siberian Times:
He has a massive nose with flared nostrils, wide open mouth, a bushy moustache and a beard. And yet all is not quite as it seems, for this sculpture, the most northerly of this genre in Asia, underwent an historic version of plastic surgery perhaps 1,500 years ago to give him a less Caucasian and more Asian appearance, according to experts. ... So the original European look of the idol was changed to a more Asian countenance. Why would this happen? 'Judging by archeological finds found inside the grottos, this anthropomorphic idol was made during the Scythian time,' Yuri Grevtsov said. 'The first change came when the more European looking face was transformed to make it appear more Mongoloid was likely to have happened in the early Middle Ages with a shift of the population in the Angara River area,' he said. In other words, incoming ethnic groups preferred the idol to be more akin to their own looks.Source: Siberia's stone idols - 2,400 year old Ust-Taseyevsky idol 'underwent racial realignment early in Middle Ages', losing his European looks by Tamara Zubchuk
Friday, October 14, 2016
Global 10: A fresh look at global genetic diversity
Update 06/02/2018: I'm working on a new version of this test. It's called the Global 25. See here.
...
Below is my new Principal Component Analysis (PCA) or genetic map of global human population structure. I think it's a little bit special, and we can discuss why in the comments if anyone's interested. The datasheet is available here; it can be used to generate 2D and 3D PCA plots, and to model samples of your choice using the nMonte and 4mix R scripts. A similar sheet with average values for the ancient populations is here.
 Here are a few examples of nMonte mixture models for highly drifted populations that often confuse the crap out of standard population genetics programs.
Here are a few examples of nMonte mixture models for highly drifted populations that often confuse the crap out of standard population genetics programs.

Karitiana Ulchi 64.45 AfontovaGora3 34.2 Dai 1.35 distance%=0.5479/distance=0.005479 Wichi Ulchi 66.8 AfontovaGora3 33.2 Dai 0 distance%=0.5055/distance=0.005055 Kalash Iran_Neolithic:I1945 38.6 Paniya 20.2 Yamnaya_Samara:I0357 17.4 Afanasievo:RISE509 16.55 Andronovo:RISE505 3.8 Iran_Late_Neolithic:I1671 3.45 Iran_Hotu:I1293 0 distance%=0.3793/distance=0.003793 Brahmin_Uttar_Pradesh Paniya 54 Yamnaya_Samara:I0357 24.55 Iran_Neolithic:I1945 21.45 Afanasievo:RISE509 0 Andronovo:RISE505 0 Iran_Hotu:I1293 0 Iran_Late_Neolithic:I1671 0 distance%=0.997/distance=0.00997 Brahmin_Tamil_Nadu Paniya 57.2 Iran_Neolithic:I1945 27.7 Yamnaya_Samara:I0357 15.1 Afanasievo:RISE509 0 Andronovo:RISE505 0 Iran_Hotu:I1293 0 Iran_Late_Neolithic:I1671 0 distance%=1.0312/distance=0.010312 Pathan Paniya 24.9 Iran_Neolithic:I1945 23.15 Andronovo:RISE505 19.95 Iran_Late_Neolithic:I1671 17.85 Yamnaya_Samara:I0357 14.15 Afanasievo:RISE509 0 Iran_Hotu:I1293 0 distance%=0.5729/distance=0.005729It's interesting to note that the Dai from southern China help to improve the fit for Karitiana from the Amazon basin, but not the Wichi from Argentina. Also, Andronovo significantly improves the fit for the East Iranian Pathans or Pashtuns, but clearly not as much for the Indo-Aryan Kalash, and none at all for Brahmins from India, who are also Indo-Aryans. Why? Don't know, but it might well be an important question in regards to the origins and spread of Indo-Iranian languages.
Monday, October 10, 2016
RIP with cannabis
Iron Age European-like people in what is now western China buried their dead with cannabis plants. Courtesy of Phys.org:
The skeleton has been identified as once belonging to a Caucasian man approximately 35 years old at the time of his death. Those that had buried him had placed a willow pillow under his head and had then placed a shroud of (13) cannabis plants over his chest reaching from below his pelvis at one end to the side of his face on the other. The skeleton lay in one of the 240 graves in the area known as the Jiayi cemetery. The people that lived in the area at the time were part of a Kingdom from 3,000 and 2,000 years ago known as the Subeixi. Prior research has shown the people lived there because it was an oasis in the desert, one that had become an important place for travelers to rest during their trek along the Silk Road.Full article: Ancient skeleton covered in cannabis shroud unearthed in China Based on this paper at Economic Botany:
Abstract: An extraordinary cache of ancient, well-preserved Cannabis plant remains was recently discovered in a tomb in the Jiayi cemetery of Turpan, NW China. Radiometric dating of this tomb and the archeobotanical remains it contained indicate that they are approximately 2800–2400 years old. Both morphological and anatomical features support the identification of the plant remains as Cannabis. Research discussed in this paper describes 13 nearly whole plants of Cannabis that appear to have been locally produced and purposefully arranged and used as a burial shroud which was placed upon a male corpse. This unique discovery provides new insight into the ritualistic use of Cannabis in prehistoric Central Eurasia. Furthermore, the fragmented infructescences of Cannabis discovered in other tombs of the Jiayi cemetery, together with similar Cannabis remains recovered from coeval tombs in the ancient Turpan cemetery along with those found in the Altai Mountains region, reveal that Cannabis was used by the local Central Eurasian people for ritual and/or medicinal purposes in the first millennium before the Christian era.Hongen Jiang et al., Ancient Cannabis Burial Shroud in a Central Eurasian Cemetery, Economic Botany (2016). DOI: 10.1007/s12231-016-9351-1 See also... Bronze Age dope dealers
Saturday, October 8, 2016
Dead cat bounce
Max Planck's Johannes Krause featured this curious map at a recent talk in Moscow on the Proto-Indo-European homeland debate (two hours into the clip here).
 It appears to be an attempt to merge several of the main competing PIE homeland hypotheses into a single "hybrid model" under the umbrella of a modified version of the recently much maligned and now practically dead Neolithic Anatolian PIE hypothesis.
Make no mistake, it is an exceedingly strange effort that will be torn apart on several levels if it ever gets published.
It appears to be an attempt to merge several of the main competing PIE homeland hypotheses into a single "hybrid model" under the umbrella of a modified version of the recently much maligned and now practically dead Neolithic Anatolian PIE hypothesis.
Make no mistake, it is an exceedingly strange effort that will be torn apart on several levels if it ever gets published.
 See also...
Big deal of 2018: Yamnaya not related to Maykop
Late PIE ground zero now obvious; location of PIE homeland still uncertain, but...
See also...
Big deal of 2018: Yamnaya not related to Maykop
Late PIE ground zero now obvious; location of PIE homeland still uncertain, but...


Tuesday, October 4, 2016
Hurrians and the others
Here's another graph based on my new D-stats datasheet. The contrast in the population affinities of Armenia_MLBA (Middle Late Bronze Age) and Armenia_EBA (Early Bronze Age) is, at least for me, surprising.
 Armenia_EBA or Kura-Araxes shows strong affinity to Caucasus populations, particularly those from the Northeast Caucasus. This is very cool, and it makes a lot of sense, because historical linguists and archaeologists generally consider Kura-Araxes people to have been early speakers of Hurrian, an ancient language thought to be closely related to present-day Northeast Caucasian languages.
But what's going on with Armenia_MLBA? I really didn't expect to see Latvians and Swedes sitting near the top of this graph. Clearly, someone from the north, closely related to present-day people from around the Baltic Sea, moved into the Armenian Plateau during or just before the Middle Bronze Age. But who were they?
I don't have a clue, but f4-stats suggest that they may have also been closely related to the Sintashta people of the Middle Bronze Age Ural steppes, who do appear very Northern European in terms of genome-wide genetic structure. The time frame fits, so does the expansive and militaristic nature of the Sintashta Culture.
Armenia_EBA or Kura-Araxes shows strong affinity to Caucasus populations, particularly those from the Northeast Caucasus. This is very cool, and it makes a lot of sense, because historical linguists and archaeologists generally consider Kura-Araxes people to have been early speakers of Hurrian, an ancient language thought to be closely related to present-day Northeast Caucasian languages.
But what's going on with Armenia_MLBA? I really didn't expect to see Latvians and Swedes sitting near the top of this graph. Clearly, someone from the north, closely related to present-day people from around the Baltic Sea, moved into the Armenian Plateau during or just before the Middle Bronze Age. But who were they?
I don't have a clue, but f4-stats suggest that they may have also been closely related to the Sintashta people of the Middle Bronze Age Ural steppes, who do appear very Northern European in terms of genome-wide genetic structure. The time frame fits, so does the expansive and militaristic nature of the Sintashta Culture.
 See also...
On Gamkrelidze-Ivanov's dubious map skills and then some
See also...
On Gamkrelidze-Ivanov's dubious map skills and then some

Yoruba Sintashta Armenia_EBA Armenia_MLBA f4 0.000676 Z 1.395 Yoruba Potapovka Armenia_EBA Armenia_MLBA f4 0.000459 Z 1.275 Yoruba Corded_Ware_CE Armenia_EBA Armenia_MLBA f4 0.000383 Z 0.933 Yoruba Poltavka Armenia_EBA Armenia_MLBA f4 0.000373 Z 0.814 Yoruba Andronovo Armenia_EBA Armenia_MLBA f4 0.000214 Z 0.486By the way, these stats are based on transversion sites only to limit the effects of post-mortem damage on the ancient samples, some of which are not UDG treated. Update 06/10/2016: As far as I can see, the qpAdm modeling software shows that Sintashta is indeed the best available proxy for the European-like admixture in Armenia_MLBA.
Outgroups Andamanese_Onge Bichon Chukchi Han Israel_Natufian Karitiana Kostenki14 MA1 Mbuti Papuan Ust_Ishim Armenia_MLBA Armenia_EBA 0.799±0.069 Sintashta 0.201±0.069 chisq 7.181 tail prob 0.618257 Armenia_MLBA Armenia_EBA 0.835±0.068 Andronovo (3) 0.165±0.068 chisq 9.549 tail prob 0.388179 Armenia_MLBA Armenia_EBA 0.842±0.065 Andronovo (4) 0.158±0.065 chisq 9.742 tail prob 0.371809 Armenia_MLBA Armenia_EBA 0.838±0.069 Srubnaya 0.162±0.069 chisq 9.993 tail prob 0.351059Update 08/10/2016: By the way, the fact that Kura-Araxes shares high genetic drift with many Indo-European-speaking Southern Europeans, such as Albanians, has no bearing on its posited identity as an Hurrian-speaking population. That's because this inflated genetic affinity is mediated via ancient groups of largely Near Eastern origin not directly related to Kura-Araxes, such as Copper and Bronze Age pre-Indo-European Europeans. On the other hand, Caucasians, particularly Northeast Caucasians, in all likelihood do share direct ancestry with Kura-Araxes.

Monday, October 3, 2016
On Gamkrelidze-Ivanov's dubious map skills and then some
I've put together a new D-stats sheet that might be useful in the Indo-European homeland debate (see here). It features new samples from the EGDP dataset, with most of the stats based on over 750K SNPs. The stats are of the form D(Chimp,Ancient)(Mbuti,X).
The idea that Indo-Iranian languages arrived in Central and South Asia from the Armenian Plateau and/or eastern Anatolia during the Bronze Age, as per Gamkrelidze and Ivanov (check out their article here and, if you don't have access, crazy map here), is still popular with a lot of people. But it's most certainly a dud.
There's too much Bronze Age steppe ancestry in this part of the world, particularly among the more isolated Indo-Iranian populations like Pamir Tajiks and the Kalasha, as well as upper caste Indians, to ignore. At the same time, there is no hard data linking any of these groups to Bronze Age Armenia or Anatolia.

 Also, some people in the comments here are still having problems comprehending the relationship between Eastern European Hunter-Gatherers (Eastern_HG or EHG) and Yamnaya, and how this relates to the issue of the Indo-European expansion from the Pontic-Caspian Steppe into other parts of Eastern Europe.
By and large the vast majority of EHG ancestry among present-day Europeans was mediated via Yamnaya or closely related groups from the steppe. How do I know? Because of the very close relationship between EHG and Yamnaya signals in present-day Europeans, and specifically Indo-European speaking Europeans, including those living in Eastern Europe.
Also, some people in the comments here are still having problems comprehending the relationship between Eastern European Hunter-Gatherers (Eastern_HG or EHG) and Yamnaya, and how this relates to the issue of the Indo-European expansion from the Pontic-Caspian Steppe into other parts of Eastern Europe.
By and large the vast majority of EHG ancestry among present-day Europeans was mediated via Yamnaya or closely related groups from the steppe. How do I know? Because of the very close relationship between EHG and Yamnaya signals in present-day Europeans, and specifically Indo-European speaking Europeans, including those living in Eastern Europe.
 Indeed, most of the EHG in Eastern Europe arrived there from the steppe as a package with Caucasus Hunter-Gatherer (Caucasus_HG or CHG) ancestry. Again, this is especially true for Indo-European speaking Northern and Eastern Europeans, who are generally hugging the line of best fit in these graphs for that reason. Note also that EHG and Yamnaya produce essentially the same results when plotted against CHG.
Indeed, most of the EHG in Eastern Europe arrived there from the steppe as a package with Caucasus Hunter-Gatherer (Caucasus_HG or CHG) ancestry. Again, this is especially true for Indo-European speaking Northern and Eastern Europeans, who are generally hugging the line of best fit in these graphs for that reason. Note also that EHG and Yamnaya produce essentially the same results when plotted against CHG.

 On the other hand, groups from far Northeastern Europe, where Uralic languages are currently spoken or were spoken until recently, appear to harbor inflated affinity to EHG and deflated affinity to CHG, putting them well above the line of best fit. These are also the same groups that show inflated East Eurasian, and more specifically Siberian, admixture, which is basically lacking in most Indo-Europeans. So I'm guessing that proto-Uralic speakers were mostly a mixture of EHG and East Eurasian, maybe with minor CHG.
Also worth noting is that Southern Europeans show inflated affinity to CHG and deflated affinity to EHG, putting them well below the line of best fit. This is because of their inflated Near Eastern ancestry (relative to Northern and Eastern Europeans) from the Neolithic and later periods, probably including CHG admixture that arrived in Southern Europe independently of EHG and Yamnaya/Yamnaya-related groups.
However, the ubiquitous presence of CHG across Europe today does not confirm Gamkrelidze and Ivanov's Armenian Plateau Indo-European homeland theory. That's because, unlike what their wacky map that I linked to above suggests, Eastern Europeans do not show any signs of ancestry from the South Caspian region (see here), which they clearly should if their Indo-European ancestors migrated en masse from Transcaucasia to the Pontic-Caspian steppe in an anti-clockwise direction around the Caspian Sea.
On the other hand, groups from far Northeastern Europe, where Uralic languages are currently spoken or were spoken until recently, appear to harbor inflated affinity to EHG and deflated affinity to CHG, putting them well above the line of best fit. These are also the same groups that show inflated East Eurasian, and more specifically Siberian, admixture, which is basically lacking in most Indo-Europeans. So I'm guessing that proto-Uralic speakers were mostly a mixture of EHG and East Eurasian, maybe with minor CHG.
Also worth noting is that Southern Europeans show inflated affinity to CHG and deflated affinity to EHG, putting them well below the line of best fit. This is because of their inflated Near Eastern ancestry (relative to Northern and Eastern Europeans) from the Neolithic and later periods, probably including CHG admixture that arrived in Southern Europe independently of EHG and Yamnaya/Yamnaya-related groups.
However, the ubiquitous presence of CHG across Europe today does not confirm Gamkrelidze and Ivanov's Armenian Plateau Indo-European homeland theory. That's because, unlike what their wacky map that I linked to above suggests, Eastern Europeans do not show any signs of ancestry from the South Caspian region (see here), which they clearly should if their Indo-European ancestors migrated en masse from Transcaucasia to the Pontic-Caspian steppe in an anti-clockwise direction around the Caspian Sea.

 Rather, it appears that CHG gene flow diffused onto the Eastern European steppe with migrants coming directly from the Caucasus (see here). And, judging by the affinities of CHG and ancient groups in large part of CHG origin, these people were more likely the speakers of Caucasian languages than of Proto-Indo-European.
Rather, it appears that CHG gene flow diffused onto the Eastern European steppe with migrants coming directly from the Caucasus (see here). And, judging by the affinities of CHG and ancient groups in large part of CHG origin, these people were more likely the speakers of Caucasian languages than of Proto-Indo-European.
 See also...
Hurrians and the others
See also...
Hurrians and the others
{kind=link}








Friday, September 30, 2016
A male-dominated conquest of Europe by Bronze Age steppe pastoralists
Just in at bioRxiv:
 Goldberg et al., Familial migration of the Neolithic contrasts massive male migration during Bronze Age in Europe inferred from ancient X chromosomes, bioRxiv, Posted September 30, 2016, doi: http://dx.doi.org/10.1101/078360
Update 15/03/2017: "Failure to Replicate a Genetic Signal for Sex Bias in the Steppe Migration into Central Europe"
Goldberg et al., Familial migration of the Neolithic contrasts massive male migration during Bronze Age in Europe inferred from ancient X chromosomes, bioRxiv, Posted September 30, 2016, doi: http://dx.doi.org/10.1101/078360
Update 15/03/2017: "Failure to Replicate a Genetic Signal for Sex Bias in the Steppe Migration into Central Europe"
Abstract: Dramatic events in human prehistory, such as the spread of agriculture to Europe from Anatolia and the Late Neolithic/Bronze Age (LNBA) migration from the Pontic-Caspian steppe, can be investigated using patterns of genetic variation among the people that lived in those times. In particular, studies of differing female and male demographic histories on the basis of ancient genomes can provide information about complexities of social structures and cultural interactions in prehistoric populations. We use a mechanistic admixture model to compare the sex-specifically-inherited X chromosome to the autosomes in 20 early Neolithic and 16 LNBA human remains. Contrary to previous hypotheses suggested by the patrilocality of many agricultural populations, we find no evidence of sex-biased admixture during the migration that spread farming across Europe during the early Neolithic. For later migrations from the Pontic steppe during the LNBA, however, we estimate a dramatic male bias, with ~5-14 migrating males for every migrating female. We find evidence of ongoing, primarily male, migration from the steppe to central Europe over a period of multiple generations, with a level of sex bias that excludes a pulse migration during a single generation. The contrasting patterns of sex-specific migration during these two migrations suggest a view of differing cultural histories in which the Neolithic transition was driven by mass migration of both males and females in roughly equal numbers, perhaps whole families, whereas the later Bronze Age migration and cultural shift were instead driven by male migration, potentially connected to new technology and conquest.

Monday, September 26, 2016
Estonian Biocentre Human Genome Diversity Panel (EGDP)
Published along with Pagani et al. 2016, the EGDP dataset is freely available at the Estonian Biocentre website as VCF and PLINK binary files here. It overlaps at ~550K SNPs with Harvard's Human Origins, and at an impressive ~1.1 million SNPs with the ~1.2 million SNP ancient DNA chip used by the Reich Lab and others.
To see what's what, I ran a Principal Component Analysis (PCA) of all of the samples except the Congo Pygmies. I then removed four Siberians that behaved as if they had very recent European ancestry, and reran the PCA. Below are a few screen grabs from the latter analysis. The datasheet is available here.


 I also tested a few of the samples with my Basal-rich K7 model. You can see their results in the spreadsheet here. Look for the individual IDs with the GS prefix.
Using the K7 spreadsheet and nMonte, here's a model for Ashkenazi Jews with some of the new EGDP populations as references, including Avars from the North Caucasus and Arabs from Israel. The Arabs do help to improve the fit, but they're not as important as Samaritans and Tuscans.
I also tested a few of the samples with my Basal-rich K7 model. You can see their results in the spreadsheet here. Look for the individual IDs with the GS prefix.
Using the K7 spreadsheet and nMonte, here's a model for Ashkenazi Jews with some of the new EGDP populations as references, including Avars from the North Caucasus and Arabs from Israel. The Arabs do help to improve the fit, but they're not as important as Samaritans and Tuscans.



Ashkenazi Anatolia_ChL 7.9 Arab_Israel_1 15.65 Avar 0.6 Bashkir 0.05 Cossack 0 Italian_Tuscan 30.45 Polish 11.75 Samaritan 33.6 Uygur 0 distance%=0.2874 / distance=0.002874In any case, that looks like a fairly sensible outcome, considering that it only took me a few minutes to put together. I've seen much worse in scientific literature.
Thursday, September 22, 2016
New rules for comments
All comments posted here must comply with the following rules. Comments that break these rules will be removed. Repeat offenders will be banned.
- no racial or ethnic taunts and insults - do not for any reason speculate about the people posting here - avoid discussions with obvious trolls and/or mentally unstable people* - no conspiracy theories - do some reading here before posting if unfamiliar with the relevant topics - do not discuss or cite any outdated physical anthropology work unless it's just the raw measurement data in the context of discussions about archeology and/or ancient DNA - do not post any illegal or defamatory material - stay on topic unless you have a good excuse for going off topic*Mental illness is not a joke, but it is a reality for a lot of people, so we're bound to see some visitors here occasionally with this problem. If unsure, simply ignore and move on. There is no rule against swearing and name calling, but try to keep things civil. Trolls should be regularly called out for who they are, but keep it short and then move on. See also... Banned commentators list
Thursday, September 15, 2016
R1a and R1b from an early Mongolian tomb
PLoS One has a new paper on a Mongolian imperial tomb dated to 1130–1250 AD, which is the early Mongolian era. Here's the table with the Y-haplogroup results:
 The authors make the sensible conclusion that these Mongolians had paternal ancestry from somewhere in Western Eurasia. But they also make this stupid comment:
The authors make the sensible conclusion that these Mongolians had paternal ancestry from somewhere in Western Eurasia. But they also make this stupid comment:


Similarly, the high frequency of R1b-M343 in geographic regions associated with the past Mongol khanates including the Golden Horde (from Ural Mountain to Western Siberia, which includes Russia, Ukraine, Belarus, Poland, Azerbaijan, Kazakhstan, and Uzbekistan), Ilkhanate (Iran and neighboring territories including Armenia, Turkey, Georgia, Afghanistan, Syria, and Tajikistan), and Chagatai Khanate (from the Aral sea to the Altai mountain, including Pakistan (Hazara), Uzbekistan, Kazakhstan, Tajikistan, India, and China), strongly suggest a close association between the Y haplotype R1b-M343 and the past Mongol Empire.No, the distribution, frequencies and subclades of R1a or R1b do not show any sort of meaningful correlation with the past Mongol Empire. The most obvious explanation, although not the only possible one, for the presence of R1a and R1b lineages in Mongolian imperial remains are the migrations of the Afanasievo and/or Andronovo people into the Altai region from the Eastern European steppes during the Bronze Age. See here...
Tuesday, September 13, 2016
ISBA7 PalaeoBarn abstracts
The abstract book for this week's meeting is available here. Emphasis is mine. The two abstracts on the genetic shifts in the East Baltic region might look as if they contradict each other, but they don't. What they're suggesting is that the East Baltic was basically home to typical European hunter-gatherers right up until the Late Neolithic, when the Corded Ware people crashed into the area, probably from the steppe via East Central Europe. For more on this topic also see here.
Mitochondrial DNA Analysis of Human Remains from Estonia Pfrengle et al. The transition from hunter-gatherer subsistence to farming is one of the most important processes in human history. In Europe, it has been found to be a result of demic diffusion originating from the Near East. The arrival of the first farmers in Europe lead to an increase of genetic diversity as well as genetic admixture of local hunter-gatherer and the migrating farmers. Previous studies investigating European human history using mitochondrial and genome-wide nuclear data from early farmers and hunter-gatherers have provided detailed insights into the process of admixture and replacement throughout the Neolithic period. However this process has been poorly studied in the Baltic region where archaeological research suggests more extensive scenarios. Here we reconstructed the complete mtDNA of 19 individuals from different archaeological sites of Estonia covering the timespan from the Narva Culture to the Corded Ware Culture and determined their mitochondrial haplogroups. The results show that the typical European hunter-gatherer maternal lineages are represented exclusively in all individuals from until the Middle Neolithic. From the Late Neolithic on, haplogroups that are associated with European Neolithic farmers are detected. The results indicate genetic continuity of foraging cultures of Mesolithic and early Neolithic backgrounds and a late demic diffusion into the territory of Estonia associated with people of the Corded Ware culture. In addition, the generated genetic data are used to gain insights into the demography of burial complexes by sex determination and maternal kinship analysis. The Neolithic Transition at the Edge of Europe Jones et al. In Europe, the Neolithic transition marked the beginning of a period of innovations which saw people move from a mobile lifestyle, dependent on hunting and gathering for survival, to a more sedentary way of life based on food production. This new lifeway, which began in the Near East ~11 kya, spread quickly across the continental interior of Europe predominantly through demic diffusion. While the genetic impact of the Neolithic transition has been well explored in central Europe, its impact on more peripheral regions of the continent has not been as extensively studied. To broaden our understanding of this dynamic phase in European prehistory, we analysed genomes from a 4,000 year temporal transect through the Baltic region spanning from the Late Mesolithic to the Late Neolithic period. We found evidence for connectivity from the Mesolithic to the Neolithic however, we also detected signals consistent with influxes from non-local populations. These influences were distinct from the early farmer admixture which transformed the genetic landscape of central Europe during the Neolithic. Interestingly, dietary stable isotope analyses (δ15N and δ 13C) show that the genetic shifts coincide with diversifications in subsistence strategy. These results suggest that the Neolithic was a period of genetic flux in the Baltic however, the cultural and technological changes observed were largely independent of forager-farmer genetic exchange. Reconstructing population history in East Asia Wang et al. The deep population history of East Asia remains poorly understood compared to that of West Eurasia, due to the lack of ancient DNA data as well as limited sampling of present-day populations especially on the Tibetan Plateau and in southern China. We report a fine scale survey of East Asian history based on genome-wide data from ancient samples in the Amur River Basin, as well as 435 newly reported individuals from 53 populations. Present-day groups can be broadly classified into highly differentiated clusters, corresponding to Amur River Basin, Tibetan Plateau, southern natives and Han Chinese. Populations of the Amur River Basin show a high degree of genetic continuity from seven thousand years ago until today, and are closely related to the strain of East Asian related ancestry present in Native Americans. Tibetan Plateau populations are all admixed, deriving about 5%-10% of their ancestry from an anciently divergent population that plausibly corresponds to the Paleolithic population on the Plateau, and the remaining part from an ancient population that no longer exists in unmixed form but that likely corresponds to expanding farmers from the Middle and Upper Yellow River Basin who also contributed 40-90% of the ancestry of Han Chinese. A total of 10-60% of Han Chinese ancestry derives from southern Native populations, and we show that the type of southern Native ancestry that contributed to Taiwan Island Austronesian speakers is most closely related to present-day speakers of Tai-Kadai languages in southern mainland China. Contextualizing the Tianyuan genome within present and ancient human genomic diversity Yang et al. Recently, many studies have produced an unprecedented number of ancient human genomes, providing insight on human dynamics in many regions, particularly West Eurasia and the Americas. Here, we present genome-wide data from the Tianyuan specimen, dating to ~40,000 years ago. Unlike other ancient genomes studied to date, the Tianyuan genome is the first ancient Upper Paleolithic sample analyzed to have contributed greatly to the East Eurasian ancestral lineage. We compare Tianyuan to several ancient and present day human genomes to better understand both the genetic diversity in the Upper Paleolithic and the similarities and differences between Tianyuan and present day populations. Overall, the addition of genome-wide Tianyuan data provides greater insight into the population history in Eurasia over the last 40,000 years. Capture of ancient genomic DNA of individuals recovered from a Medieval Alemannic gravesite provides evidence for high mobility of fellowships during the 7th century CE. O'Sullivan et al. Whether the historic spread of cultural/language groups such as the Alemanni were migrations or local adoption of culture is still unresolved in archaeology. The Alemanni were a confederation of tribes that inhabited an area, from the third to the 10th century CE, which approximately overlaps with the modern distribution of Alemannic German dialect in Swabia. We present the genomic and isotopic data of eight individuals excavated from a gravesite in Niederstotzingen, Germany of supposed Alemannic origin dated to the 7th century CE. There were two multiple burials at the site suggesting either kinship or fellowship between the individuals. The tombs in the gravesite contained cultural artefacts and weapons indicating close contact of the Alemanni with Longobards and Byzantines. We investigated the genetic affinity of these individuals between each other and to modern West Eurasians. The genetic analysis utilised the targeted enrichment and sequencing of over 1.2 million genetic markers that have known ascertainment. From these data, we found no familial relationship among the individuals in the multiple graves, thus supporting a burial practice based rather on fellowship. All individuals were genetically male. The genetic affinities of the individuals, based on modern genetic distributions, were five Eastern Europeans, two Germans/Austrians and one Southern European. Isotopic data supports that only the Southern European individual was certainly born outside this region. The genetic data appear to correlate with the provenance of the burial artefacts, showing that westward movements and interactions among cultural groups likely occurred in this region during the 7th century CE. Origins and genetic legacy of the first people in Remote Oceania Skoglund et al. The appearance of people associated with the Lapita culture in the South Pacific ~3,000 years ago marked the beginning of the last major human dispersal to unpopulated lands, culminating in the settlement of eastern Polynesia ~1,000-700 years ago. However, the genetic relationship of these pioneers to the long established Papuan peoples of the New Guinea region is debated. We report the first genome-wide ancient DNA data from Asia-Pacific region, from four ~2,900 to ~2,500 year old Lapita culture individuals from Vanuatu and Tonga, and co-analyze them with new data from 356 present-day Oceanians. Today, all indigenous people of the South Pacific harbor a mixture of ancestry from Papuans and a population of East Asian origin that we find to be a statistical match to the ancient Lapita individuals. Most analyses have interpreted the ubiquitous Papuan ancestry in the region today-at least 25%-as evidence that the first humans to reach Remote Oceania and Polynesia were derived from mixtures near New Guinea prior to the Lapita expansion into Remote Oceania. Our results refute this scenario, as none of the geographically and temporally diverse Lapita individuals had detectable Papuan ancestry. These results imply later major human population movements, which spread Papuan ancestry through the South Pacific after the islands' first peopling. An ancient genomic perspective on the horse domestication process Librado et al. The domestication of the horse in the Pontic-Caspian steppes some 6,000 years ago represents one major turning point in human history. With horses, humans could travel for the first time well above their own speed and carry their germs, culture and genes across vast geographic areas. The development of horse-drawn chariots and cavalry also radically changed the history of warfare and was instrumental to the emergence of transcontinental empires. Additionally, beyond the battlefield, farm horses have massively impacted agricultural productivity. The biological changes that accompanied the process of horse domestication are, however, difficult to reconstruct from current patterns of genetic diversity both due to the development of intensively selected and extremely influential breeds during the last two centuries, and the almost extinction of wild horses. Recent developments in ancient DNA research have opened for the characterization of complete genomes, epigenomes and microbiota over long time series. We have applied such approaches to a large panel of horse remains spread across Eurasia and dated to 44,000-200 years ago. This started revealing the genetic structure of horse populations prior to and during early domestication stages as well as the history of genetic changes that accompanied their further transformation in a range of cultural contexts. I will present our latest progress made on an extensive dataset of ancient horse genomes spanning the whole domestication temporal and geographical range. Mobility between the Aegean and the Levant in the Late Second Millennium BCE: inference from ancient DNA of pigs Meiri et al. The Late Bronze and the early Iron Ages (ca. 1450-950 BCE) of the eastern Mediterranean region are characterized by dramatic historical processes. Empires emerged and collapsed, trade connections were established and severed, and at the end of this era socio-political unrest and migration of large groups of people were rife throughout the region. In the 12th century BCE the movements of the so-called "Sea Peoples" affected wide parts of the East Mediterranean. We study the nature of human movements during this period on trade connections, culture and animal husbandry using the ancient DNA of domestic animals, above all pigs. We recently showed that in Israel, European pig haplotypes appeared ca. 900 BCE, and soon after took over the gene pool, with all modern wild boars in Israel carrying European mitochondrial DNA. Here, we broadened the chronological and spatial scopes by studying ancient pig mitochondrial DNA from the southern Levant and Greece. The Near Eastern haplotype Y1 and supposedly Near Eastern haplotype Y2 were discovered in Greece in the mid to late 3rd millennium BCE, while the European haplotypes were found in Israel in the early Iron Age IIA (ca. 900 BCE). We propose that pigs were moved between Europe and Anatolia since the early Bronze Age. Connections between Greece and the southern Levant are observed in the Iron Age, and probably result from the migration of Sea Peoples to the east. These results shed light on networks and movements of people during both times of prosperity and crisis.
Brachycephalization on the western steppe after the Mesolithic
Interesting stuff from the ADNABIOARC team at Scientific Reports today:

Abstract: The Neolithic transition brought about fundamental social, dietary and behavioural changes in human populations, which, in turn, impacted skeletal morphology. Crania are shaped through diverse genetic, ontogenetic and environmental factors, reflecting various elements of an individual’s life. To determine the transition’s effect on cranial morphology, we investigated its potential impact on the face and vault, two elements potentially responding to different influences. Three datasets from geographically distant regions (Ukraine, Iberia, and the Levant plus Anatolia) were analysed. Craniometric measurements were used to compare the morphology of pre-transition populations with that of agricultural populations. The Neolithic transition corresponds to a statistically significant increase only in cranial breadth of the Ukrainian vaults, while facial morphology shows no consistent transformations, despite expected changes related to the modification of masticatory behaviour. The broadening of Ukrainian vaults may be attributable to dietary and/or social changes. However, the lack of change observed in the other geographical regions and the lack of consistent change in facial morphology are surprising. Although the transition from foraging to farming is a process that took place repeatedly across the globe, different characteristics of transitions seem responsible for idiosyncratic responses in cranial morphology.Cheronet et al., Morphological change in cranial shape following the transition to agriculture across western Eurasia, Scientific Reports, Published online: 13 September 2016, doi:10.1038/srep33316 See also.. Modeling Steppe_EMBA
Thursday, September 8, 2016
ASHG 2016 abstracts
The American Society of Human Genetics (ASHG) 2016 Meeting Online Planner and Abstract Search can be accessed here. Unless I'm missing something, there's surprisingly little ancient genomics stuff this year considering the huge advances in this area recently.
However, Iain Mathieson has an interesting contribution about interactions between Balkan foragers and early Balkan farmers. Reading between the lines, I reckon we can probably expect some R1b in these Balkan ancients. This shouldn't be surprising, however, considering the publication earlier this year of the Ice Age Villabruna genome from present-day northeast Italy, which belonged to R1b and, as pointed out by myself, also showed strong genome-wide ties to ancient Siberians (see here).
Genome-wide ancient DNA from Europe’s first encounter of farmers and hunter-gatherers. I. Mathieson.
Oleg Balanovsky has been acting a little strange lately; seemingly going out of his way to fight the Kurgan Indo-European paradigm in various papers and talks. In this poster he underlines the fact that the Yamnaya people from the North Caspian region did not totally vacate their homeland nor moved en masse to Western Europe, but actually left most of their descendants in the North Caspian. OK Oleg, but almost all modern day Europeans, especially those from across Northern and Eastern Europe, do derive a lot of their ancestry from Bronze Age steppe groups closely related to the Yamnaya people of the North Caspian. This will become even more obvious soon with the publication of new ancient data from the Baltic region.
Y-chromosomal sequencing and screening reveal both stability and migrations in North Eurasian populations. O. Balanovsky.
And here we have what appears to be yet another hilariously wrong paper in the making on the population history of South Asia. Why do they mention Anatolia? How about the Arctic Circle? Enough already. Get some goddamn ancient genomes from South Asia and finally do it right.
Genetic variation reveals migrations into the Indian subcontinent and its influence on the Indian society. A. Bose, D.E. Platt, L. Parida, P. Paschou, P. Drineas.
These abstracts also sparked my interest, in a positive way:
Using whole-genome sequencing to shed insight on the complex prehistory of Sardinia. C. Chiang
Y-chromosomal composition of mediaeval and contemporary populations in Norway and adjacent Scandinavian countries: Y-STR haplotypes and the rare Y-haplogroup Q. B. Berger.
A complex history of archaic admixture in modern humans. R. Bohlender, Y. Yu, C. Huff, A. Rogers.
Genomic insights into the population structure and history of the Irish Travellers. E.H. Gilbert.
Friday, September 2, 2016
Ust'-Ishim man x2
Just wanted to see what would happen if I split Ust-Ishim into two homozygous sequences, and ran him in a Principal Component Analysis (PCA) alongside other fully homozygous individuals and composites, both modern day and ancient. In theory this shouldn't be a problem, considering that his genome is of such an incredibly high quality (~42-fold coverage). The datasheets for the PCA below are available here and here.
This is the only way that I can think of to run a PCA in which genetic drift specific to the Ust'-Ishim population of ~45K YBP western Siberia has a clear impact. Indeed, lots of interesting things on these plots. Note, for instance, that on the second plot the Afanasievo and Yamnaya-Catacomb composites appear to be attracted to the Caucasus Hunter-Gatherer composite. Also note how unusual the Neolithic Iranians (Iran_EN) appear.



 Update 02/09/2016: Interactive 3D versions of these PCA are now available at Open Genomes here and here. Below are screen caps of angles that appear to recapitulate the geography of Eurasia. See comments below for details.
Update 02/09/2016: Interactive 3D versions of these PCA are now available at Open Genomes here and here. Below are screen caps of angles that appear to recapitulate the geography of Eurasia. See comments below for details.

 See also...
Ancient vs modern day West Eurasian variation
See also...
Ancient vs modern day West Eurasian variation
Subscribe to:
Posts (Atom)